Recombinant Mouse Beta-arrestin-2 (Arrb2)
-
货号:CSB-YP835595MO
-
规格:
-
来源:Yeast
-
其他:
-
货号:CSB-EP835595MO
-
规格:
-
来源:E.coli
-
其他:
-
货号:CSB-EP835595MO-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
货号:CSB-BP835595MO
-
规格:
-
来源:Baculovirus
-
其他:
-
货号:CSB-MP835595MO
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
别名:Arrb2Beta-arrestin-2; Arrestin beta-2
-
种属:Mus musculus (Mouse)
-
蛋白长度:full length protein
-
表达区域:1-410
-
氨基酸序列MGEKPGTRVF KKSSPNCKLT VYLGKRDFVD HLDKVDPVDG VVLVDPDYLK DRKVFVTLTC AFRYGREDLD VLGLSFRKDL FIATYQAFPP MPNPPRPPTR LQDRLLKKLG QHAHPFFFTI PQNLPCSVTL QPGPEDTGKA CGVDFEIRAF CAKSIEEKSH KRNSVRLIIR KVQFAPETPG PQPSAETTRH FLMSDRRSLH LEASLDKELY YHGEPLNVNV HVTNNSAKTV KKIRVSVRQY ADICLFSTAQ YKCPVAQLEQ DDQVSPSSTF CKVYTITPLL SDNREKRGLA LDGQLKHEDT NLASSTIVKE GANKEVLGIL VSYRVKVKLV VSRGGDVSVE LPFVLMHPKP HDHITLPRPQ SAPRETDVPV DTNLIEFDTN YATDDDIVFE DFARLRLKGM KDDDCDDQFC
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Functions in regulating agonist-mediated G-protein coupled receptor (GPCR) signaling by mediating both receptor desensitization and resensitization processes. During homologous desensitization, beta-arrestins bind to the GPRK-phosphorylated receptor and sterically preclude its coupling to the cognate G-protein; the binding appears to require additional receptor determinants exposed only in the active receptor conformation. The beta-arrestins target many receptors for internalization by acting as endocytic adapters (CLASPs, clathrin-associated sorting proteins) and recruiting the GPRCs to the adapter protein 2 complex 2 (AP-2) in clathrin-coated pits (CCPs). However, the extent of beta-arrestin involvement appears to vary significantly depending on the receptor, agonist and cell type. Internalized arrestin-receptor complexes traffic to intracellular endosomes, where they remain uncoupled from G-proteins. Two different modes of arrestin-mediated internalization occur. Class A receptors, like ADRB2, OPRM1, ENDRA, D1AR and ADRA1B dissociate from beta-arrestin at or near the plasma membrane and undergo rapid recycling. Class B receptors, like AVPR2, AGTR1, NTSR1, TRHR and TACR1 internalize as a complex with arrestin and traffic with it to endosomal vesicles, presumably as desensitized receptors, for extended periods of time. Receptor resensitization then requires that receptor-bound arrestin is removed so that the receptor can be dephosphorylated and returned to the plasma membrane. Mediates endocytosis of CCR7 following ligation of CCL19 but not CCL21. Involved in internalization of P2RY1, P2RY4, P2RY6 and P2RY11 and ATP-stimulated internalization of P2RY2. Involved in phosphorylation-dependent internalization of OPRD1 and subsequent recycling or degradation. Involved in ubiquitination of IGF1R. Beta-arrestins function as multivalent adapter proteins that can switch the GPCR from a G-protein signaling mode that transmits short-lived signals from the plasma membrane via small molecule second messengers and ion channels to a beta-arrestin signaling mode that transmits a distinct set of signals that are initiated as the receptor internalizes and transits the intracellular compartment. Acts as signaling scaffold for MAPK pathways such as MAPK1/3 (ERK1/2) and MAPK10 (JNK3). ERK1/2 and JNK3 activated by the beta-arrestin scaffold are largely excluded from the nucleus and confined to cytoplasmic locations such as endocytic vesicles, also called beta-arrestin signalosomes. Acts as signaling scaffold for the AKT1 pathway. GPCRs for which the beta-arrestin-mediated signaling relies on both ARRB1 and ARRB2 (codependent regulation) include ADRB2, F2RL1 and PTH1R. For some GPCRs the beta-arrestin-mediated signaling relies on either ARRB1 or ARRB2 and is inhibited by the other respective beta-arrestin form (reciprocal regulation). Increases ERK1/2 signaling in AGTR1- and AVPR2-mediated activation (reciprocal regulation). Involved in CCR7-mediated ERK1/2 signaling involving ligand CCL19. Is involved in type-1A angiotensin II receptor/AGTR1-mediated ERK activity. Is involved in type-1A angiotensin II receptor/AGTR1-mediated MAPK10 activity. Is involved in dopamine-stimulated AKT1 activity in the striatum by disrupting the association of AKT1 with its negative regulator PP2A. Involved in AGTR1-mediated chemotaxis. Appears to function as signaling scaffold involved in regulation of MIP-1-beta-stimulated CCR5-dependent chemotaxis. Involved in attenuation of NF-kappa-B-dependent transcription in response to GPCR or cytokine stimulation by interacting with and stabilizing CHUK. Suppresses UV-induced NF-kappa-B-dependent activation by interacting with CHUK. The function is promoted by stimulation of ADRB2 and dephosphorylation of ARRB2. Involved in IL8-mediated granule release in neutrophils. Involved in p53/TP53-mediated apoptosis by regulating MDM2 and reducing the MDM2-mediated degradation of p53/TP53. May serve as nuclear messenger for GPCRs. Upon stimulation of OR1D2, may be involved in regulation of gene expression during the early processes of fertilization. Also involved in regulation of receptors other than GPCRs. Involved in endocytosis of TGFBR2 and TGFBR3 and down-regulates TGF-beta signaling such as NF-kappa-B activation. Involved in endocytosis of low-density lipoprotein receptor/LDLR. Involved in endocytosis of smoothened homolog/Smo, which also requires GRK2. Involved in endocytosis of SLC9A5. Involved in endocytosis of ENG and subsequent TGF-beta-mediated ERK activation and migration of epithelial cells. Involved in Toll-like receptor and IL-1 receptor signaling through the interaction with TRAF6 which prevents TRAF6 autoubiquitination and oligomerization required for activation of NF-kappa-B and JUN. Involved in insulin resistance by acting as insulin-induced signaling scaffold for SRC, AKT1 and INSR. Involved in regulation of inhibitory signaling of natural killer cells by recruiting PTPN6 and PTPN11 to KIR2DL1. Involved in the internalization of the atypical chemokine receptor ACKR3. Acts as an adapter protein coupling FFAR4 receptor to specific downstream signaling pathways, as well as mediating receptor endocytosis. During the activation step of NLRP3 inflammasome, directly associates with NLRP3 leading to inhibition of proinflammatory cytokine release and inhibition of inflammation.
-
基因功能参考文献:
- Data show that inactivation of the beta-arrestin-2 gene, barr2, in beta-cells of adult mice greatly impairs insulin release and glucose tolerance . PMID: 28145434
- Spinal Arrb2 may serve as an intracellular gate for acute to chronic pain transition via desensitization of NMDAR. PMID: 27538456
- Knockout of mice beta-arrestin2, but not beta-arrestin1, results in deficits in plasticity mediated by mGlu1 receptors in CA3 pyramidal neurons and by mGlu5 receptors in CA1 pyramidal neurons. PMID: 27886171
- beta-arrestin 2 can protect liver tissue from LPS-induced injury via inhibition of TLR4/NF-kappaB signaling pathway-mediated inflammation. PMID: 29375207
- Non-visual arrestins regulate the focal adhesion formation via small GTPases RhoA and Rac1 independently of G-protein-coupled receptors. PMID: 29133163
- Arrb2 induces cardiomyocyte death by interacting with the p85 subunit of PI3K, and negatively regulating the formation of p85-PI3K/CaV3 survival complex, thus blocking activation of PI3K-Akt-GSK3beta cell survival signalling pathway in cardiac ischemia-reperfusion. PMID: 29016703
- these results suggest that beta-arrestin2 down-regulation increases hepatocellular carcinoma cell migration and invasion ability PMID: 27759077
- beta-Arrestin2 increases the Itch-mediated ubiquitylation of SuFu PMID: 29515120
- Arrestin beta2 deficiency-mediated intestinal progenitor/stem cell radioprotection relied on protracted NF-kappaB activation and subsequent suppression of PUMA induction. PMID: 27128598
- GIP stimulation induces a switch in GIPR recycling from a rapid endosomal to a slow trans-Golgi network (TGN) pathway. GPCR kinases and b-arrestin2 are required for this switch in recycling. PMID: 27974210
- selective inactivation of the GPCR-associated protein beta-arrestin 2 in hepatocytes of adult mice results in greatly increased hepatic GCGR signaling, leading to striking deficits in glucose homeostasis PMID: 28650340
- AT1R-beta-arrestin-2 pathway signaling plays an important role in renal fibrosis. PMID: 28274926
- These data suggest that one allele of arrestin-2 is unable to support normal locomotor behavior due to signaling and/or developmental defects. PMID: 28419873
- Beta-arrestin-2 with beta-arrestin-1 shared common mechanisms to suppress podocyte autophagy by negative regulation of ATG12-ATG5 conjugation. PMID: 27054338
- [beta]-arrestin2 regulates intestinal mucosal inflammation under both homeostatic and colitic conditions. Its mode of action involves negative regulation of T-cell activation and its requirement for induction of regulatory T cells. PMID: 26296063
- Results suggest that the antipruritic effects of kappa opioid receptor agonists may not require betaarrestin2 PMID: 26318102
- This shows that mood stabilizers lamotrigine, lithium and valproate can exert behavioral effects in mice by disrupting the beta-arrestin 2-mediated regulation of Akt/GSK3 signaling by D2 dopamine receptors. PMID: 26459714
- findings show for the first time that Ang II receptor signaling to beta-arrestin regulates ARF6 activation. These proteins together control receptor endocytosis and ultimately cell migration. PMID: 26703465
- These results reveal that the protective effect of deficiency of Arrb2 is due to loss of negative regulation of Akt. PMID: 26582201
- that Insulin-like growth factor-1 contributes to the mucosal repair by beta-arrestin2-mediated extracellular signal-regulated kinase signaling in experimental colitis PMID: 26362717
- ARRB2 is not involved in hepatocellular carcinogenesis. PMID: 26077142
- morphine activated JNK2 through an arrestin-independent Src- and PKC-dependent mechanism, whereas fentanyl activated JNK2 through a Src-GRK3/arrestin-2-dependent and PKC-independent mechanism. PMID: 26056051
- Arrb2 is required for potentiation of odorant receptor responses by Chrm3. PMID: 25800153
- Beta arrestin 2 participates in the perpetuation of airway hyperresponsiveness in a murine asthma model. PMID: 25569510
- GPR40 functions via both G protein-mediated and beta-arrestin-mediated mechanisms; endogenous and synthetic ligands differentially engage these pathways to promote insulin secretion. PMID: 26157145
- Arrb2 knockout prevents development of cellular mu opioid receptor tolerance but does not affect opioid-withdrawal-related adaptations in single periaqueductal grey neurons PMID: 24597632
- The anti-inflammatory action exerted by beta-arrestin-2 appears to be mediated in part through the direct inhibition of p38MAPK, preventing NF-kB activation, and in part through cAMP and PKA activation primed by G protein signaling. PMID: 25318610
- beta-arrestin-biased signaling regulates memory reconsolidation PMID: 25831532
- findings suggest specific localization of beta-arrestin2 in the myenteric plexus within MOR1-expressing neurons and provide a relation for direct intracellular crosstalk between activation and beta-arrestin2 signaling in the myenteric neurons. PMID: 25083714
- ARRB2 is unlikely to be involved in the regulation of insulin secretion, but it is required for beta cell mass plasticity. PMID: 24317793
- behavioral studies indicate that GPCR-driven beta-arrestin2 sequestration plays an important peripheral role in the development of thermal sensitivity PMID: 24695785
- These results reveal a novel mechanism by which beta-arrestin 2 negatively regulates TLR4-mediated inflammatory reactions PMID: 25012660
- Identify N-methyl-d-aspartate receptor as down-regulator of inflammasome activity via beta-arrestin-2 pathway. PMID: 25104498
- a novel adaptor beta-arrestin 2 that associates with phosphorylated TIGIT for further recruitment of SHIP1 (SH2-containing inositol phosphatase 1) through the ITT-like motif. PMID: 24817116
- results support a model in which beta-arrestin 2 inhibits the production of proinflammatory chemokines, which limits the recruitment of myeloid immune cells and thereby attenuates allergic skin inflammation PMID: 24577407
- Beta-arrestin2 regulates cannabinoid CB1 receptors in brain. PMID: 24094141
- In Ghrhr D60G homozygous and heterozygous pbARR2-Cre, PTEN mice. PMID: 23689346
- These results suggest that beta-arrestin-2 negatively regulates alcohol preference and reward PMID: 23839051
- Loss of beta-arrestin 2 reduces the number of Foxp3+ CD4+ regulatory T cells, exacerbating autoimmune encephalomyelitis. PMID: 23859136
- beta-arrestin2 plays a protective role in myocardial inflammation-induced inflammation. PMID: 23861891
- CCR1.beta-arrestin-2 complex may be related to a potential scavenging function of the receptor, which may be important for maintenance of chemokine gradients and receptor responsiveness in complex fields of chemokines during inflammation. PMID: 24056371
- morphine suppresses TLR4-induced TNF release in mast cells, preventing the IKK-dependent phosphorylation of SNAP-23, which is necessary for TNF exocytosis, and this inhibition correlates with the formation of a beta-arrestin-2/TRAF6 complex PMID: 23960234
- our results reveal a negative regulatory role for beta-arr2 in polymicrobial infection-induced inflammation PMID: 23753627
- beta-arrestin2 deficient pancreatic islets exhibit blunted insulin secretion. The number of docked insulin granules in beta-arrestin2 deficient islets is decreased compared to wild-type islets, while insulin content and beta cell mass remains unchanged. PMID: 23660189
- Suggest beta-arrestin-2-dependent signaling plays a major role in AngII-induced abdominal aortic aneurysm formation. PMID: 23524589
- beta-arrestin 2 physically associates with the Aph-1a subunit of the gqamma-secretase complex and redistributes the complex toward detergent-resistant membranes, increasing the catalytic activity of the complex. PMID: 23202293
- Deletion of GSK3beta in D2R-expressing neurons reveals distinct roles for beta-arrestin2 signaling in antipsychotic and lithium action. PMID: 23188793
- The Cftr/betaarr2 double knockout mice also exhibit wild-type (WT) levels of cholesterol synthesis, and WT profiles of signaling protein expression have previously been shown to be altered in CF due to cholesterol-related pathways. PMID: 22523395
- Heterologous (but not homologous) desensitization required beta-arrestin-2 (betaarr-2). PMID: 22689562
- Data suggest that the proteinase-activated receptor-2 PAR(2)-enhanced inflammatory process is beta-arrestin-2 dependent, whereas the protective anticonstrictor effect of bronchial epithelial PAR(2) may be beta-arrestin independent. PMID: 23012429
显示更多
收起更多
-
亚细胞定位:Cytoplasm. Nucleus. Cell membrane. Membrane, clathrin-coated pit. Cytoplasmic vesicle. Note=Translocates to the plasma membrane and colocalizes with antagonist-stimulated GPCRs.
-
蛋白家族:Arrestin family
-
组织特异性:Predominantly localized in neuronal tissues and in the spleen.
-
数据库链接:
KEGG: mmu:216869
STRING: 10090.ENSMUSP00000099623
UniGene: Mm.203747
Most popular with customers
-

Recombinant Human Intestinal-type alkaline phosphatase (ALPI) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
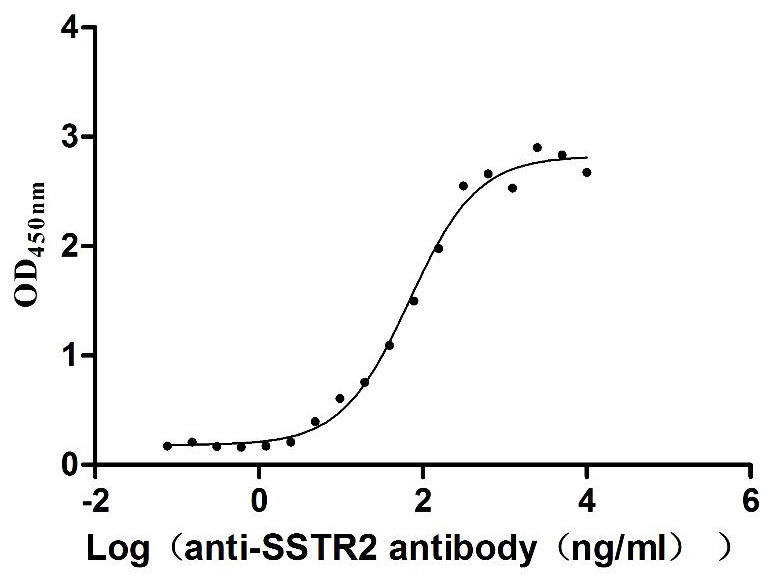
Recombinant Human Somatostatin receptor type 2 (SSTR2)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
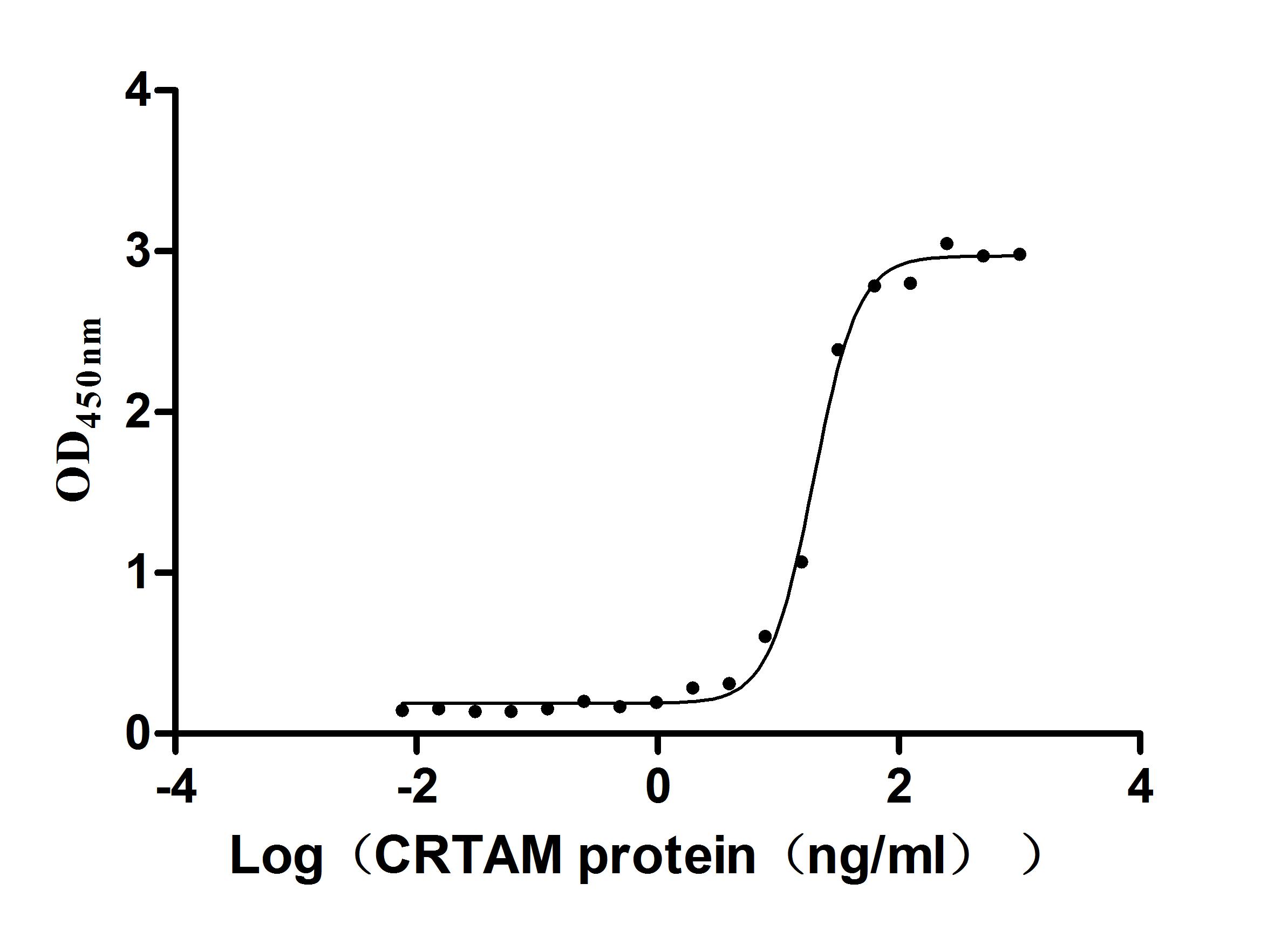
Recombinant Human Cell adhesion molecule 1 (CADM1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
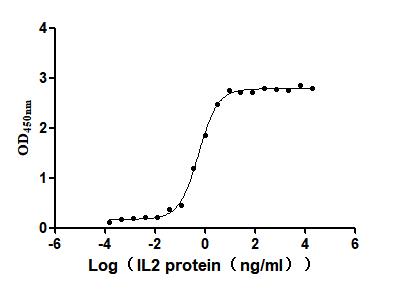
Recombinant Human Interleukin-2 (IL2) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 8(CEACAM8) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human B- and T-lymphocyte attenuator(BTLA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
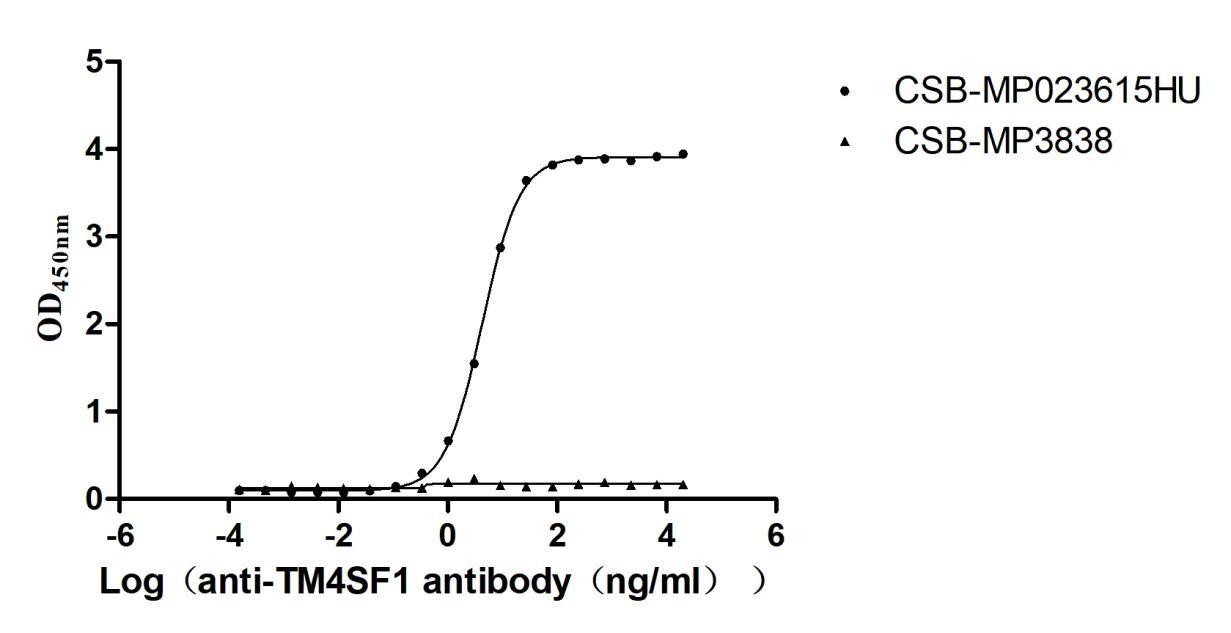
Recombinant Human Transmembrane 4 L6 family member 1(TM4SF1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Gastric inhibitory polypeptide receptor(GIPR),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)










