osk Antibody
-
货号:CSB-PA333049XA01DLU
-
规格:¥440
-
促销:

-
图片:
-
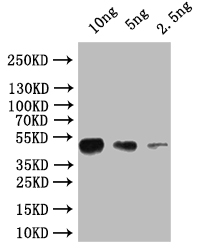 Western Blot
Western Blot
Positive WB detected in: recombinant protein
All lanes: osk Antibody at 1:1000
Secondary
Goat polyclonal to rabbit IgG at 1/50000 dilution
Predicted band size: 45 kDa
Observed band size: 45 kDa
-
-
其他:
产品详情
-
产品名称:Rabbit anti-Drosophila melanogaster (Fruit fly) osk Polyclonal antibody
-
Uniprot No.:P25158
-
基因名:osk
-
别名:Maternal effect protein oskar osk CG10901
-
宿主:Rabbit
-
反应种属:Drosophila melanogaster (Fruit fly)
-
免疫原:Recombinant Drosophila melanogaster (Fruit fly) osk protein (1-260aa)
-
免疫原种属:Drosophila melanogaster (Fruit fly)
-
标记方式:Non-conjugated
-
克隆类型:Polyclonal
-
抗体亚型:IgG
-
纯化方式:>95%, Protein G purified
-
浓度:It differs from different batches. Please contact us to confirm it.
-
保存缓冲液:Preservative: 0.03% Proclin 300
Constituents: 50% Glycerol, 0.01M PBS, pH 7.4 -
产品提供形式:Liquid
-
应用范围:ELISA, WB
-
推荐稀释比:
Application Recommended Dilution WB 1:500-1:5000 -
Protocols:
-
储存条件:Upon receipt, store at -20°C or -80°C. Avoid repeated freeze.
-
货期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time maybe differs from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
产品评价

相关产品
靶点详情
-
功能:Organizes the germ plasm and directs localization of the posterior determinant nanos. Oskar protein is required to keep nos RNA and staufen protein at the posterior pole.
-
基因功能参考文献:
- A missense mutation in the dynactin Arp1 subunit causes most oskar mRNA to localise in the posterior cytoplasm rather than cortically. PMID: 29035202
- oskar transport signals are weak by necessity; their weakness facilitates transfer of the oskar mRNA from the oocyte transport machinery to the machinery for posterior localization. PMID: 28760927
- The long isoform of the protein Oskar regulates the maternal inheritance of mitochondria. Long Oskar traps and maintains mitochondria at the posterior at the site of primordial germ cell (PGC) formation through an actin-dependent mechanism. Mutating long oskar strongly reduces the number of mtDNA molecules inherited by PGCs. Therefore, Long Oskar ensures germline transmission of mitochondria to the next generation. PMID: 27923120
- Crystal structure identifies OSK domain as RNA-binding domain and LOTUS domain as Vasa-binding domain. PMID: 26190108
- Osk has a role in the regulation of stability, regulation of translation, and localization of relevant mRNAs through direct interaction with their 3'UTRs PMID: 26324911
- The study determined the SOLE structure formed after alternative splicing of Oskar mRNA as an helical structure with few noncanonical base pairs, capped by a pentanucleotide loop that seems could accommodate a protein partner. PMID: 26089324
- results therefore suggest that efficient posterior localization of oskar mRNA requires the concerted activities of both Dynein and Kinesin-1. PMID: 24244700
- These results reveal that Par-1 controls the timing of pole plasm assembly by promoting the localization of oskar mRNA but inhibiting the accumulation of Short Oskar protein. PMID: 23948254
- Oskar protein was required for recruiting EB1 and CLIP-190 to the oocyte posterior; posterior enrichment of EB1 and CLIP-190 is necessary for high levels of endocytosis in this region of the cell; therefore a functional link between dynamic oocyte microtubules and endocytosis. PMID: 22561189
- The study demonstrates that the exon junction complex associates with oskar mRNA upon splicing in vitro and that Drosophila exon junction complex deposition is constitutive and conserved. PMID: 22426546
- report that a Golgi-endosomal protein, Mon2, acts downstream of Osk to remodel cortical actin and to anchor the pole plasm PMID: 21610029
- osk was independently lost in multiple holometabolous insect lineages and that these losses are phylogenetically correlated with changes in germline determination strategies in these species. PMID: 21552321
- Translational repression is mediated by BREs, regulatory elements positioned in two clusters near both ends of the oskar mRNA 3' UTR. PMID: 20230756
- Data indicate that posteriorly localized Par-1 regulates posterior patterning by stabilizing Osk. PMID: 11951092
- Data show that cortical anchoring of the posterior determinant Oskar is a crucial step in pole plasm assembly and restriction, required for proper development of Drosophila melanogaster. PMID: 12117819
- Kinesin restricts pole plasm formation to the posterior by moving oskar mRNA away from microtubule-rich lateral and anterior cortical regions. PMID: 12134163
- MOESIN crosslinks actin and cell membrane in Drosophila oocytes and is required for anchoring of this protein. PMID: 12477397
- Data suggest that Orb-mediated cytoplasmic polyadenylation stimulates oskar translation to achieve the high levels of Oskar protein necessary for posterior patterning and germline differentiation. PMID: 12538512
- We conclude that Bruno plays similar roles in translational regulation of gurken and oskar PMID: 12591598
- Cup is a translational repressor of oskar that is required to assemble the oskar mRNA localization machinery. We propose that Cup coordinates localization with translation. PMID: 14691132
- splicing at the first exon-exon junction of oskar RNA is essential for oskar mRNA localization at the posterior pole PMID: 15118729
- Hrp48 colocalizes with oskar mRNA throughout oogenesis, and interacts with its 5' and 3' regulatory regions, suggesting that it binds directly to oskar mRNA to mediate its posterior transport. PMID: 15130488
- The mechanisms that prevent accumulation of Oskar protein until it can be secured at the posterior pole of the oocyte include regulated degradation or inhibition of translational elongation. PMID: 15239960
- The vlsnull mutant shows that valois+ is required for high levels of Oskar protein to accumulate during oogenesis PMID: 15634703
- link between oskar translation control and localization in oogenesis PMID: 16715044
- oskar RNA acts as a scaffold or regulatory RNA essential for development of the oocyte. PMID: 16835436
- Oskar allows nanos mRNA translation in Drosophila embryos by preventing its deadenylation by Smaug/CCR4. PMID: 17050620
- An oskar-dependent positive feedback loop maintains the polarity of the Drosophila oocyte. PMID: 17275299
- A late phase in accumulation of Osk protein, typically not monitored because of imperviousness of late stage oocytes to antibodies, is crucial for body patterning. PMID: 17359300
- We propose that Oskar maintains its localization at the posterior pole through dual functions in regulating endocytosis and F-actin dynamics. PMID: 17419993
- Osk stimulates endosomal cycling, which in turn promotes F-actin reorganization to anchor the pole plasm components to the oocyte cortex. PMID: 18272590
- By following oskar mRNA particles in living oocytes, study shows that the mRNA is actively transported along microtubules in all directions, with a slight bias toward the posterior. PMID: 18775316
- We observe oskar mRNA to oligomerize as hundreds of copies forming large particles which are necessary for its long range transport and localization. We show the formation of these particles occurs in the nurse cell nucleus in an Hrp48-dependent manner PMID: 19597554
显示更多
收起更多
-
组织特异性:Begins to accumulate at the posterior pole of the oocyte from stage 8 onwards.
-
数据库链接:
KEGG: dme:Dmel_CG10901
STRING: 7227.FBpp0081435
UniGene: Dm.7184
Most popular with customers
-
-
YWHAB Recombinant Monoclonal Antibody
Applications: ELISA, WB, IF, FC
Species Reactivity: Human, Mouse, Rat
-
Phospho-YAP1 (S127) Recombinant Monoclonal Antibody
Applications: ELISA, WB, IHC
Species Reactivity: Human
-
-
-
-
-