Recombinant Mouse Aryl hydrocarbon receptor nuclear translocator-like protein 1 (Arntl), partial
-
中文名称:Recombinant Mouse Aryl hydrocarbon receptor nuclear translocator-like protein 1(Arntl),partial
-
货号:CSB-EP895734MO1f3
-
规格:¥2328
-
图片:
-
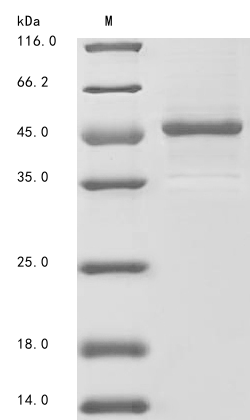 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
产品详情
-
纯度:Greater than 85% as determined by SDS-PAGE.
-
基因名:Arntl
-
Uniprot No.:
-
别名:Arntl; Bmal1Aryl hydrocarbon receptor nuclear translocator-like protein 1; Arnt3; Brain and muscle ARNT-like 1
-
种属:Mus musculus (Mouse)
-
蛋白长度:Partial of Isoform 3
-
来源:E.coli
-
分子量:46.5 kDa
-
表达区域:49-434aa
-
氨基酸序列EYAEHQGRIKNAREAHSQIEKRRRDKMNSFIDELASLVPTCNAMSRKLDKLTVLRMAVQHMKTLRGATNPYTEANYKPTFLSDDELKHLILRAADGFLFVVGCDRGKILFVSESVFKILNYSQNDLIGQSLFDYLHPKDIAKVKEQLSSSDTAPRERLIDAKTGLPVKTDITPGPSRLCSGARRSFFCRMKCNRPSVKVEDKDFASTCSKKKADRKSFCTIHSTGYLKSWPPTKMGLDEDNEPDNEGCNLSCLVAIGRLHSHMVPQPANGEIRVKSMEYVSRHAIDGKFVFVDQRATAILAYLPQELLGTSCYEYFHQDDIGHLAECHRQVLQTREKITTNCYKFKIKDGSFITLRSRWFSFMNPWTKEVEYIVSTNTVVLANVLE
Note: The complete sequence including tag sequence, target protein sequence and linker sequence could be provided upon request. -
蛋白标签:C-terminal 6xHis-Myc-tagged
-
产品提供形式:Liquid or Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
缓冲液:Tris-based buffer,50% glycerol
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
产品评价

相关产品
靶点详情
-
功能:Transcriptional activator which forms a core component of the circadian clock. The circadian clock, an internal time-keeping system, regulates various physiological processes through the generation of approximately 24 hour circadian rhythms in gene expression, which are translated into rhythms in metabolism and behavior. It is derived from the Latin roots 'circa' (about) and 'diem' (day) and acts as an important regulator of a wide array of physiological functions including metabolism, sleep, body temperature, blood pressure, endocrine, immune, cardiovascular, and renal function. Consists of two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system. Both the central and peripheral clocks can be reset by environmental cues, also known as Zeitgebers (German for 'timegivers'). The predominant Zeitgeber for the central clock is light, which is sensed by retina and signals directly to the SCN. The central clock entrains the peripheral clocks through neuronal and hormonal signals, body temperature and feeding-related cues, aligning all clocks with the external light/dark cycle. Circadian rhythms allow an organism to achieve temporal homeostasis with its environment at the molecular level by regulating gene expression to create a peak of protein expression once every 24 hours to control when a particular physiological process is most active with respect to the solar day. Transcription and translation of core clock components (CLOCK, NPAS2, ARNTL/BMAL1, ARNTL2/BMAL2, PER1, PER2, PER3, CRY1 and CRY2) plays a critical role in rhythm generation, whereas delays imposed by post-translational modifications (PTMs) are important for determining the period (tau) of the rhythms (tau refers to the period of a rhythm and is the length, in time, of one complete cycle). A diurnal rhythm is synchronized with the day/night cycle, while the ultradian and infradian rhythms have a period shorter and longer than 24 hours, respectively. Disruptions in the circadian rhythms contribute to the pathology of cardiovascular diseases, cancer, metabolic syndromes and aging. A transcription/translation feedback loop (TTFL) forms the core of the molecular circadian clock mechanism. Transcription factors, CLOCK or NPAS2 and ARNTL/BMAL1 or ARNTL2/BMAL2, form the positive limb of the feedback loop, act in the form of a heterodimer and activate the transcription of core clock genes and clock-controlled genes (involved in key metabolic processes), harboring E-box elements (5'-CACGTG-3') within their promoters. The core clock genes: PER1/2/3 and CRY1/2 which are transcriptional repressors form the negative limb of the feedback loop and interact with the CLOCK|NPAS2-ARNTL/BMAL1|ARNTL2/BMAL2 heterodimer inhibiting its activity and thereby negatively regulating their own expression. This heterodimer also activates nuclear receptors NR1D1/2 and RORA/B/G, which form a second feedback loop and which activate and repress ARNTL/BMAL1 transcription, respectively. ARNTL/BMAL1 positively regulates myogenesis and negatively regulates adipogenesis via the transcriptional control of the genes of the canonical Wnt signaling pathway. Plays a role in normal pancreatic beta-cell function; regulates glucose-stimulated insulin secretion via the regulation of antioxidant genes NFE2L2/NRF2 and its targets SESN2, PRDX3, CCLC and CCLM. Negatively regulates the mTORC1 signaling pathway; regulates the expression of MTOR and DEPTOR. Controls diurnal oscillations of Ly6C inflammatory monocytes; rhythmic recruitment of the PRC2 complex imparts diurnal variation to chemokine expression that is necessary to sustain Ly6C monocyte rhythms. Regulates the expression of HSD3B2, STAR, PTGS2, CYP11A1, CYP19A1 and LHCGR in the ovary and also the genes involved in hair growth. Plays an important role in adult hippocampal neurogenesis by regulating the timely entry of neural stem/progenitor cells (NSPCs) into the cell cycle and the number of cell divisions that take place prior to cell-cycle exit. Regulates the circadian expression of CIART and KLF11. The CLOCK-ARNTL/BMAL1 heterodimer regulates the circadian expression of SERPINE1/PAI1, VWF, B3, CCRN4L/NOC, NAMPT, DBP, MYOD1, PPARGC1A, PPARGC1B, SIRT1, GYS2, F7, NGFR, GNRHR, BHLHE40/DEC1, ATF4, MTA1, KLF10 and also genes implicated in glucose and lipid metabolism. Promotes rhythmic chromatin opening, regulating the DNA accessibility of other transcription factors. May play a role in spermatogenesis; contributes to the chromatoid body assembly and physiology. The NPAS2-ARNTL/BMAL1 heterodimer positively regulates the expression of MAOA, F7 and LDHA and modulates the circadian rhythm of daytime contrast sensitivity by regulating the rhythmic expression of adenylate cyclase type 1 (ADCY1) in the retina. The preferred binding motif for the CLOCK-ARNTL/BMAL1 heterodimer is 5'-CACGTGA-3', which contains a flanking Ala residue in addition to the canonical 6-nucleotide E-box sequence. CLOCK specifically binds to the half-site 5'-CAC-3', while ARNTL binds to the half-site 5'-GTGA-3'. The CLOCK-ARNTL/BMAL1 heterodimer also recognizes the non-canonical E-box motifs 5'-AACGTGA-3' and 5'-CATGTGA-3'. Essential for the rhythmic interaction of CLOCK with ASS1 and plays a critical role in positively regulating CLOCK-mediated acetylation of ASS1. Plays a role in protecting against lethal sepsis by limiting the expression of immune checkpoint protein CD274 in macrophages in a PKM2-dependent manner. Regulates the diurnal rhythms of skeletal muscle metabolism via transcriptional activation of genes promoting triglyceride synthesis (DGAT2) and metabolic efficiency (COQ10B).
-
基因功能参考文献:
- BMAL1 Deficiency Contributes to Mandibular Dysplasia by Upregulating MMP3. PMID: 29276151
- Data suggest that loss of aryl hydrocarbon receptor nuclear translocator-like protein (BMAL1) function in astrocytes results in altered GABA levels, leading to the over-inhibition of the circuits involved in learning and memory and to the uncoupling the suprachiasmatic nucleus (SCN) oscillators. PMID: 28186121
- Circadian clock protein BMAL1 regulates IL-1beta in macrophages via NRF2. PMID: 30127006
- The expression of the clock gene Bmal1 in myeloid cells regulates the innate and adaptive immune responses that mediate autoimmune diseases. PMID: 29234010
- Insulin reduces Bmal1 transcriptional activity by affecting its intracellular localization in mouse liver, which requires Akt-dependent phosphorylation of the Ser42 residue. PMID: 27576939
- Bmal1 modulates lipoprotein production and biliary cholesterol excretion by regulating the expression of Mtp and Abcg5/Abcg8 via Shp and Gata4. PMID: 27721414
- these results suggest a role for bmal1 in the development of asthmatic airway disease via the regulation of lung antiviral responses to common viral triggers of asthma PMID: 28401936
- The daily rhythm of BMAL1 mRNA was completely abolished in the EGR1-deficient mice. PMID: 29138967
- Clock:BMAL1 promotes a transcriptionally permissive chromatin landscape that primes its target genes for transcription activation rather than directly activating transcription. Environmental or pathological conditions can reprogram the rhythmic expression of clock-controlled genes. PMID: 29300726
- Bmal1 and Dio2 are required to maintain cone photoreceptor functional integrity. PMID: 29045837
- Islet cell maturation was also characterized by induction in the expression of circadian transcription factor BMAL1, deletion of which altered postnatal development of glucose-stimulated insulin secretion and the associated transcriptional network. PMID: 29500314
- PML mediates the binding of PER2 to BMAL1 in the BMAL1/CLOCK heterodimer and is an important component in the organization of a functional clock complex in the nucleus. PMID: 27383066
- The results demonstrate that Bmal1 expression in skeletal muscle is both necessary and sufficient to regulate total sleep amount and reveal that critical components of normal sleep regulation occur in muscle. PMID: 28726633
- The findings indicate that BMAL1 deregulation is a feature of the mTOR-activated state and suggest a molecular mechanism for mitigating circadian phenotypes in a neurodevelopmental disorder. PMID: 28746872
- Data show that CRY1 binds directly to the PAS domain core of CLOCK:BMAL1, driven primarily by interaction with the CLOCK PAS-B domain. PMID: 28143926
- Data suggest that BMAL1 protein expression in the SCN (suprachiasmatic nucleus) is higher in night-time compared to day-time in both chow-fed and fcHFHS-fed mice; the same pattern is found in the LHb (epithalamic lateral habenula) for BMAL1 expression, being higher at night. (fcHFHS = free choice high-fat high-sugar diet) PMID: 28687372
- PTEN ablation led to downregulation of BMI-1, a critical regulator of adult stem cell self-renewal, and elevated senescence, suggesting the presence of a protective system that prevents transformation. Short- and long-term PTEN depletion followed by activated BMAL1, a core clock protein, contributed to accumulation of hair follicle stem cells. PMID: 28602615
- These findings indicate that the expressions of two key clock genes, Per1 and Bmal1, in the SCN are regulated in such a way that they may adopt different phases and free-running periods relative to each other and are respectively associated with the expression of activity onset and offset. PMID: 28416676
- Expression of Per2 and Bmal1 in the ovary did not display clear diurnal oscillation. PMID: 28213822
- The Smad3 and Bmal1 regulate p21 and S100A4 expression in myocardial stromal fibroblasts through TNF-alpha. PMID: 28721450
- Data demonstrated that the circadian rhythm of testosterone synthesis in TM3 cells was disturbed following Fen treatment as evidenced by changes in the circadian rhythmicity of core clock genes (Bmal1, Rev-erbalpha, Roralpha). PMID: 29040059
- TFEB regulates PER3 expression via glucose-dependent effects on CLOCK/BMAL1 PMID: 27373683
- These data suggest that forebrain clock timing plays a critical role in shaping the efficiency of learning and memory retrieval over the circadian day. PMID: 27091299
- The circadian clock component Bmal1 is involved in blood brain barrier homeostasis. PMID: 28912161
- Study shows a role for Bmal1, the endogenous circadian clock, in glucose metabolism in the skeletal muscle. Findings implicate altered molecular clock dictating significant changes in altered substrate metabolism in the absence of feeding or activity changes. PMID: 27486508
- A significant proportion of circadian lncRNAs are expressed at enhancer regions, mostly bound by two key circadian transcription factors, BMAL1 and REV-ERBalpha. PMID: 28335007
- A cis/trans isomerization switch in the BMAL1 transactivation domain controls circadian rhythms. Conformationally locking the switch or inhibiting prolyl isomerases influences circadian period to suggest that slow protein dynamics play a role in timekeeping. PMID: 28506462
- Bmal1 deficiency in macrophages promotes atherosclerosis by enhancing recruitment of Ly6chi monocytes to atherosclerotic lesions. PMID: 27927724
- Bmal1 autophagic degradation and Bmal1 role in glucose homeostasis PMID: 26228022
- These results suggest that miR-27b-3p plays an important role in the posttranscriptional regulation of BMAL1 protein in the liver. PMID: 26919869
- Bmal1 is a potent negative regulator in myeloid cells in the context of allergic asthma. PMID: 28336811
- this study shows that BMAL1 can regulate cellular innate immunity against specific RNA viruses PMID: 27913791
- Data demonstrate that CLOCK and BMAL1 regulate muscle insulin sensitivity via SIRT1, and activation of SIRT1 might be a potential valuable strategy to attenuate muscle insulin resistance related to circadian misalignment. PMID: 27035655
- Bmal1 role in oocyte fertilization, early embryo development and implantation PMID: 27140828
- neural signaling in select behavioral centers is impaired in the absence of BMAL1, likely underlying Bmal1 KO male copulatory defects, demonstrating the importance of the BMAL1 protein in the maintenance of neural circuits that drive pheromone-mediated mating behaviors. PMID: 27704948
- Using circular chromosome conformation capture sequencing, we systematically examined the interacting loci of a Bmal1-bound super-enhancer upstream of a clock gene Nr1d1 in mouse liver. Global analysis showed that cohesin-CTCF co-binding sites tend to insulate the phases of circadian oscillating genes while cohesin-non-CTCF sites are associated with high circadian rhythmicity of transcription. PMID: 27135601
- Renal circadian clocks control a variety of metabolic/homeostatic processes at the intrarenal and systemic levels and are involved in drug disposition PMID: 27056296
- Data show a dissociation of Molecular and Endocrine Circadian Rhythms in Male Mice Lacking Bmal1 in the Adrenal Cortex PMID: 27690690
- Report altered BMAL1/CLOCK expression in conditions of chronically altered blood flow. PMID: 27923220
- DNA-binding mode for the CLOCK:BMAL1 heterotetramer complex PMID: 28414715
- results demonstrate that Bmal1 promotes osteoblast differentiation by regulating BMP2 expression in MC3T3-E1 cells PMID: 27506892
- Studies indicate the broad and complex actions of brain and muscle-ARNT-like factor (Bmal1/BMAL1) on physiology and behavior. PMID: 27916643
- this paper shows abnormal collagen VI expression in Bmal1-/- mice, a model for circadian rhythm deregulation PMID: 26945058
- expression of oscillatory genes was dampened in the adult-life Bmal1 knockout mice, whereas overall gene expression was largely unchanged. PMID: 26843191
- Bmal1 is required for beta cell compensatory expansion, survival and metabolic adaptation to diet-induced obesity in mice PMID: 26762333
- BMAL1 deficiency results in a low bone mass phenotype. PMID: 26789548
- Loss of Bmal1 solely from adult skeletal muscle results in reductions in specific tension, increased oxidative fibre type and increased muscle fibrosis . iMSBmal1-/- mice develop increased bone calcification and decreased joint collagen. PMID: 26486627
- The amplitude of the day-night fluctuation of Bmal1 expression was reduced in the amygdala and hippocampus of light-dark-shifted mice. PMID: 26498235
- circadian rhythmicity of ghrelin signaling requires Bmal1 and is driven by a food-responsive clock in the gastric ghrelin-secreting cell that not only regulates ghrelin, but also GOAT activity. PMID: 26576661
- Both clock- and feeding-dependent mechanisms regulate the daily accumulation of key enzymes in polyamine biosynthesis through rhythmic binding of BMAL1:CLOCK to conserved DNA elements PMID: 26456331
显示更多
收起更多
-
亚细胞定位:Nucleus. Cytoplasm. Nucleus, PML body.
-
组织特异性:Expressed in liver and testis (at protein level). Expressed in the suprachiasmatic nucleus (SCN) in a circadian manner.
-
数据库链接:
KEGG: mmu:11865
STRING: 10090.ENSMUSP00000046235
UniGene: Mm.33970
Most popular with customers
-
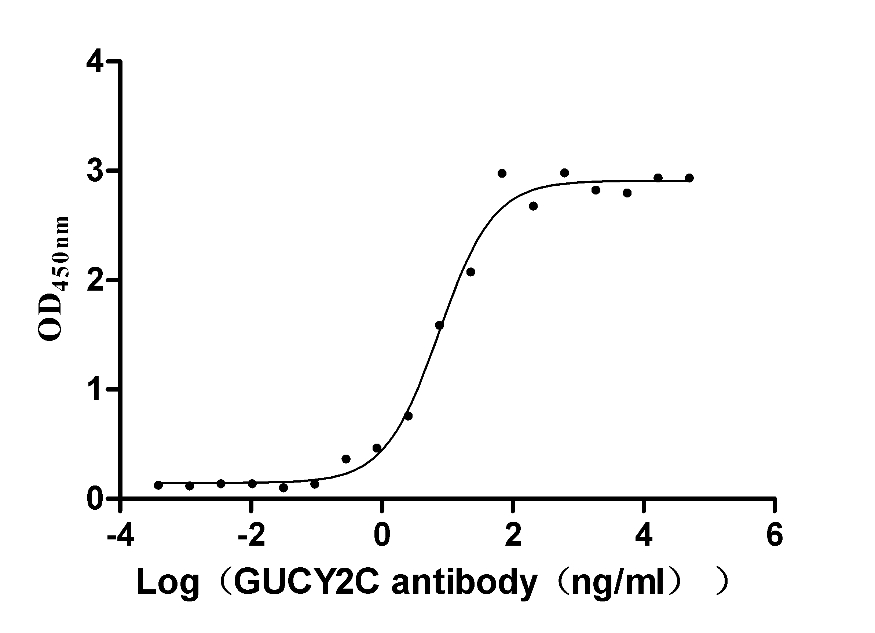
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
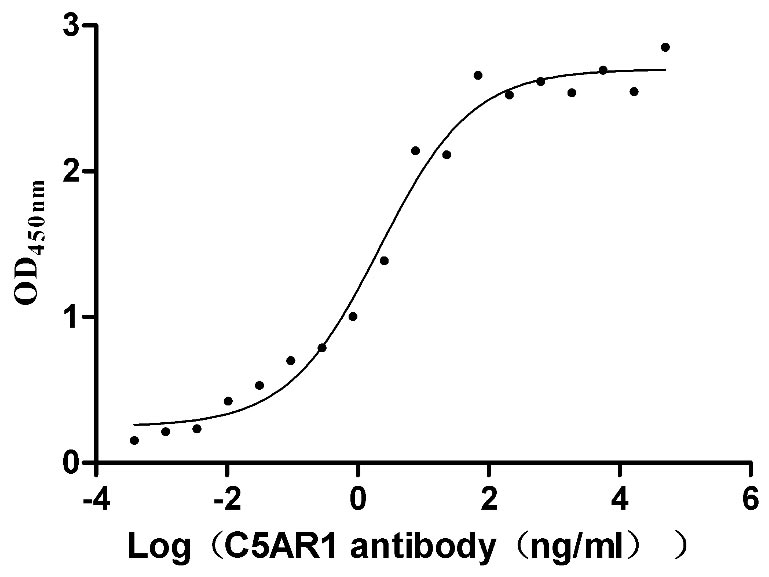
Recombinant Human C5a anaphylatoxin chemotactic receptor 1 (C5AR1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Mouse Transthyretin (Ttr) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Mouse Tyrosine-protein kinase Mer (Mertk), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)










