Recombinant Mouse Patatin-like phospholipase domain-containing protein 2 (Pnpla2), partial
-
中文名称:小鼠Pnpla2重组蛋白
-
货号:CSB-YP803885MO
-
规格:
-
来源:Yeast
-
其他:
-
中文名称:小鼠Pnpla2重组蛋白
-
货号:CSB-EP803885MO
-
规格:
-
来源:E.coli
-
其他:
-
中文名称:小鼠Pnpla2重组蛋白
-
货号:CSB-EP803885MO-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名称:小鼠Pnpla2重组蛋白
-
货号:CSB-BP803885MO
-
规格:
-
来源:Baculovirus
-
其他:
-
中文名称:小鼠Pnpla2重组蛋白
-
货号:CSB-MP803885MO
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:Pnpla2
-
Uniprot No.:
-
别名:Pnpla2; Atgl; Patatin-like phospholipase domain-containing protein 2; EC 3.1.1.3; Adipose triglyceride lipase; Desnutrin
-
种属:Mus musculus (Mouse)
-
蛋白长度:Partial
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Catalyzes the initial step in triglyceride hydrolysis in adipocyte and non-adipocyte lipid droplets. Exhibits a strong preference for the hydrolysis of long-chain fatty acid esters at the sn-2 position of the glycerol backbone. Also has acylglycerol transacylase activity. Acts coordinately with LIPE/HLS and DGAT2 within the lipolytic cascade. Regulates adiposome size and may be involved in the degradation of adiposomes. May play an important role in energy homeostasis. May play a role in the response of the organism to starvation, enhancing hydrolysis of triglycerides and providing free fatty acids to other tissues to be oxidized in situations of energy depletion.
-
基因功能参考文献:
- ATGL in brown adipose tissue is not a prerequisite for cold-induced thermogenesis in vivo. PMID: 28988821
- Non-cardiac ATGL-mediated modulation of the cardiac lipidome may play an important role in the pathogenesis of chronic heart failure. PMID: 29320510
- this study shows that long non-coding steroid receptor RNA activator promotes hepatic steatosis through repressing the expression of ATGL PMID: 27759039
- ATGL deficiency results in male infertility in mice. PMID: 28510703
- Loss of ATGL induces spontaneous development of pulmonary neoplasia in knockout mouse model. PMID: 27213586
- ATGL acts as an important upstream signaling node that, via SIRT1, increases autophagy/lipophagy as a means to promote hepatic lipid droplet catabolism. PMID: 28380348
- CGI-58 regulates hepatic neutral lipid storage and inflammation in the genetic absence of ATGL. PMID: 27396333
- Enhanced lipolysis in response to mitochondrial uncoupling relies on a form of autophagy as lipid droplets are captured by endolysosomal vesicles which is HSL/ATGL-independent. PMID: 28488768
- The Atgl is down-regulated by the basal transcription factor Sp1 in preadipocytes and that the magnitude of down-regulation depends on interactions between Sp1 and peroxisome proliferator-activated receptor gamma (PPARgamma). PMID: 28726642
- TAG synthesis and levels of PUFA-TAGs were lowered by the diacylglycerol acyltransferase (DGAT)1 inhibitor, T863. The lipase inhibitor, Atglistatin, increased the levels of TAG in both WT and ATGL-deficient mouse Hepatic stellate cell (HSC). Both Atglistatin and T863 inhibited the induction of activation marker, alpha-smooth muscle actin, in rat HSCs, but not in mouse HSCs. PMID: 27179362
- G0S2 protein but not mRNA levels were reduced in the adipose tissue of ATGL-deficient mice, corroborating the involvement of ATGL in the stabilization of G0S2 PMID: 27248498
- Atgl deficiency induces podocyte apoptosis and leads to glomerular filtration barrier damage. PMID: 28194887
- These data raise the possibility that ATGL deficiency could impair the renal fatty acid metabolism though inhibiting PPARalphaexpression, which may lead to lipid deposition and cell apoptosis of PCT, and finally contribute to the renal fibrosis and dysfunction PMID: 28400046
- Markers of mitochondrial content and respiration were increased in adipose tissue from ATGL knockout mice. PMID: 27357546
- ase. Similar changes of GPNMB and G0S2 expression were present in a human liposarcoma database. These results show that a previously-unknown, fully penetrant epistatic interaction between Pnpla2 and Lipe can cause liposarcoma in mice. DAKO mice provide a promising model for studying early premalignant changes that lead to late-onset malignant disease. PMID: 28459858
- ANGPTL8 is a stress-response protein that down-regulates expression of ATGL. PMID: 26569053
- ATGL KO mice had significantly lower fasting plasma free fatty-acids compared to WT. Fasting endogenous glucose production and peripheral glucose metabolism were similar in KO and WT mice. PMID: 26236747
- Pharmacological inhibition of ATGL may prove useful to prevent HFD-induced obesity and insulin resistance PMID: 26508640
- A role for Atgl in kidney lipolysis:fasting up-regulates Atgl levels and phosphorylation in mouse kidney. PMID: 25879679
- data provide mechanistic insights into Jordans' anomaly in neutrophils and suggest that ATGL is a potent regulator of immune cell function and inflammatory diseases. PMID: 26109679
- Data show that obese adipocyte-specific adipose triglyceride lipase (ATGL) knockout (AAKO) mice had reduced adipocyte lipolysis. PMID: 26196542
- transcriptional transactivation assays with a luciferase reporter system revealed that Fabps enhance the ability of Atgl/Cgi-58-mediated lipolysis to induce the activity of peroxisome proliferator-activated receptors. PMID: 25953897
- Data (including data from studies in atherosclerotic knockout mice) suggest CGI58- (abhydrolase domain containing 5-)deficient macrophages store-up triglyceride droplets and have reduced phagocytic capacity, comparable to Atgl-deficient macrophages. PMID: 25316883
- Data suggest that cardiotrophin-1 up-regulates lipolysis in white adipocytes via 1) induction of perilipin, 2) activation of hormone sensitive lipase (via phosphorylation by PKA), and 3) inactivation of ATGL (via up-regulation of its inhibitor G0S2). PMID: 25351614
- 4E-BP1/2-null cells express less ATGL and accumulate more fat than control cells, while knock down of Egr1 in 4E-BP1/2-null cells increases ATGL expression and decreases fat storage. PMID: 25814662
- ATGL acts as retinyl ester hydrolase in hepatic stellate cells promoting the degradation of retinyl ester stores in addition to its established function as Triglyceride lipase. PMID: 25732851
- ATGL overexpression in C2C12 myotubes increased lipolysis, which was associated with increased peroxisome proliferator-activated receptor (PPAR)- partial differential activity, transcriptional upregulation of some PPAR partial differential target genes, and enhanced mitochondrial capacity. PMID: 25852007
- Adipocyte rather than skeletal myocyte ATGL-mediated lipolysis plays a greater role during acute exercise in part because of compensatory mechanisms that maintain lipolysis in muscle, but not adipose tissue, when ATGL is absent. PMID: 25783895
- The anti-lipolytic activity of the HCV core protein alters ATGL binding to CGI-58 and enhances the association of both proteins with lipid droplets. PMID: 25381252
- L-FABP is not required to channel ATGL-hydrolyzed FAs to mitochondria for beta-oxidation or the nucleus for PPAR-alpha regulation PMID: 24610891
- ESR1 regulates ATGL and perilipin-mediated lipid metabolism and droplet size in femurs from mice. PMID: 23296636
- Data (including data from studies using site-directed mutagenesis of ATGL/PNPLA2) suggest threonine-372 is phosphorylation site that plays critical role in regulating subcellular distribution and lipolytic activity of ATGL/PNPLA2 in white adipocytes. PMID: 24801391
- ATGL promotes the expression of genes related to mitochondrial oxidative metabolism (activate the PPARa-PGC-1a complex). PMID: 24096872
- Desnutrin-catalyzed lipolysis activates PPAR delta, rather than PPAR alpha, which in turn is critical for expression of genes involved in mitochondrial function, and thus ATP production, required for glucose-stimulated insulin secretion. PMID: 24268737
- Systemic knockout of adipose triglyceride lipase (ATGL), the pivotal enzyme of triglyceride lipolysis, results in a murine phenotype that is characterized by progredient cardiac steatosis and severe heart failure PMID: 24657704
- These findings unravel a novel protective role of ATGL against hepatic inflammation which could have important implications for metabolic and inflammatory liver diseases. PMID: 24002947
- acquired cardiomyocyte-specific ATGL deficiency is sufficient to promote fibrotic and hypertrophic cardiomyopathy and impair myocardial fatty acid oxidation in the absence of markedly reduced PPARalpha signalling PMID: 23708736
- It added genetic stress due to knockout (KO) of adipocyte triglyceride lipase (ATGL), the rate limiting enzyme in lipid droplets(LD) lipolysis. PMID: 23740967
- These results suggest that reduced ATGL and CGI-58 expression and increased TG synthesis may exacerbate myocardial steatosis and oxidative stress, thereby promoting cardiac apoptosis in diabetic mice. PMID: 23886955
- chronic reduction in myocardial TAG content by cardiomyocyte specific ATGL overexpression is able to prevent doxorubicin induced cardiac dysfunction. PMID: 23704323
- data suggest that ATGL(-/-)mSMC have a distinct phenotype that may be related to vascular pathogenesis PMID: 23583398
- Data suggest that increased expression of Atgl in myocardium is adaptive response to accumulation of myocardial triglycerides in diabetes; study included mice with either heterozygous ATGL deficiency or cardiomyocyte-specific ATGL overexpression. PMID: 23349479
- the role of ATGL in modulating PPARalpha-dependent processes also in the small intestine PMID: 23220585
- FSP27 acts to constitutively limit the lipid droplet presence of ATGL, perilipin 1 plays an essential role in mediating the response of ATGL action to beta-adrenergic hormones. PMID: 23204327
- functional muscle lipolysis depends on both CGI-58 and ATGL PMID: 23413028
- data suggest that hepatic ATGL knockdown enhances glucose tolerance by increasing hepatic glucose utilization and uncouples impairments in insulin action from hepatic TAG accumulation PMID: 22993196
- analysis ofsubstrate and stereo/regioselectivity of adipose triglyceride lipase, hormone-sensitive lipase, and diacylglycerol-O-acyltransferases PMID: 23066022
- Studies suggest that beta-adrenergic activation can result in PKA-mediated phosphorylation of ATGL Ser(406), to moderately increase ATGL-mediated lipolysis. PMID: 22733971
- investigation of role of Atgl in skeletal muscle of Atgl knockout (KO) mice: Mitochondrial oxidative phosphorylation is unaltered in Atgl KO mice. Electrostimulation yields reduced force production/higher relaxation in Atgl KO mice. PMID: 22496349
- genetic control of ATGL-mediated lipolysis modulates adipose triglyceride level and resistance to obesity and diabetes PMID: 22383686
显示更多
收起更多
-
亚细胞定位:Lipid droplet. Cell membrane; Single-pass type II membrane protein.
-
组织特异性:Expressed at high levels in white and brown adipose tissue, and to a lesser degree in testis and cardiac muscle. Barely detected in liver, spleen, thymus, kidney, skeletal muscle, and brain. Among the white adipose depots, gonadal fat showed the highest l
-
数据库链接:
KEGG: mmu:66853
STRING: 10090.ENSMUSP00000127149
UniGene: Mm.29998
Most popular with customers
-
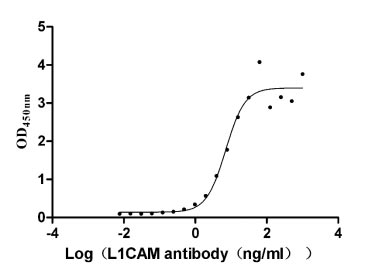
Recombinant Human Neural cell adhesion molecule L1 (L1CAM), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
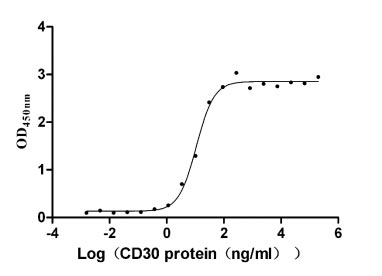
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human Claudin-6 (CLDN6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
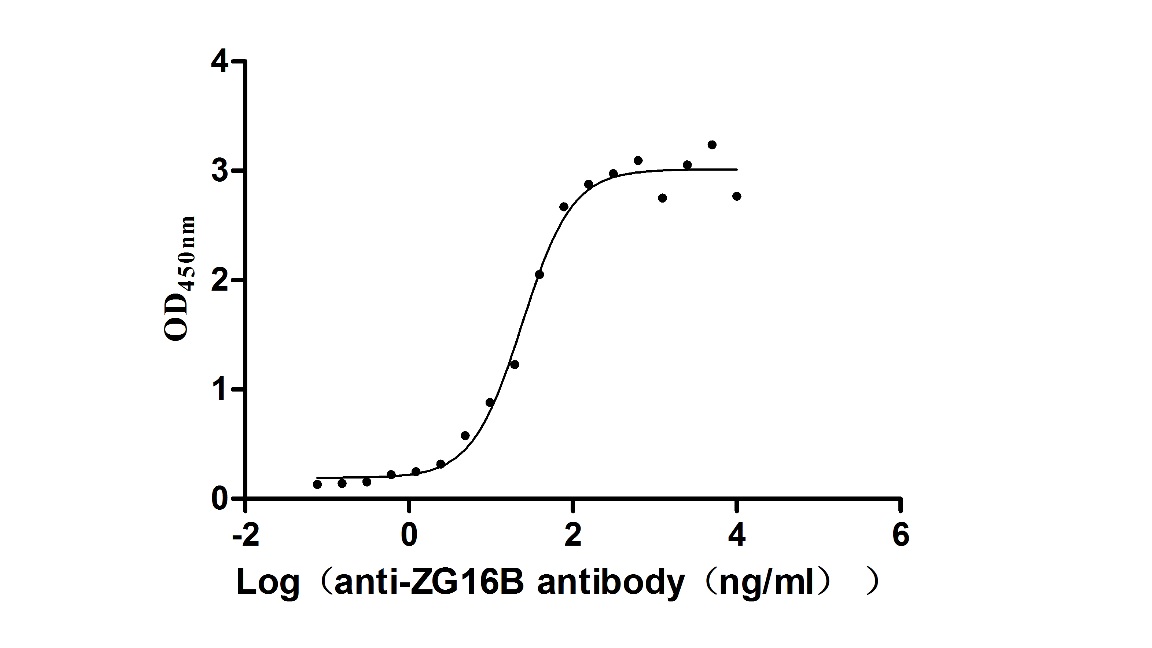
Recombinant Macaca fascicularis zymogen granule protein 16 homolog B (ZG16B) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
-
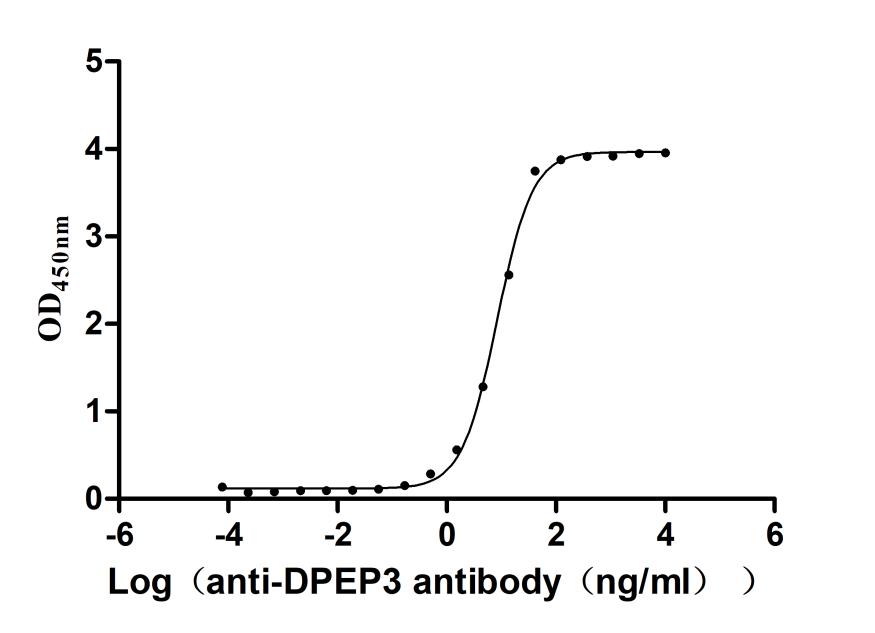
Recombinant Macaca fascicularis Dipeptidase 3(DPEP3) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
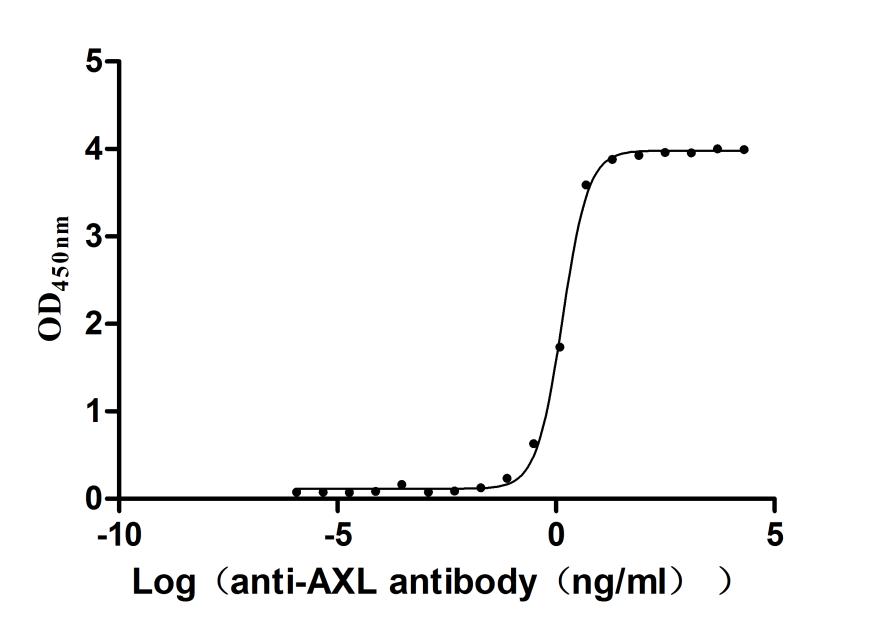
Recombinant Human Tyrosine-protein kinase receptor UFO(AXL),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)