细胞因子受体
大家知道,细胞因子一般是通过与细胞表面相应的细胞因子受体结合而发挥生物学作用。细胞因子与其受体结合后,会启动复杂的细胞内分子相互作用,最终引起细胞基因转录的变化。
已知的细胞因子受体绝大多数是跨膜蛋白,由胞外、跨膜和胞质区组成。胞外膜区是识别结合细胞因子的部位,胞质区在受体激活后启动信号转导。
根据细胞因子受体的结构,可分为不同的家族或超家族,包括免疫球蛋白(Ig)超家族、I型细胞因子受体、II型细胞因子受体、肿瘤坏死因子受体(TNFR)超家族和趋化因子受体(图1)。
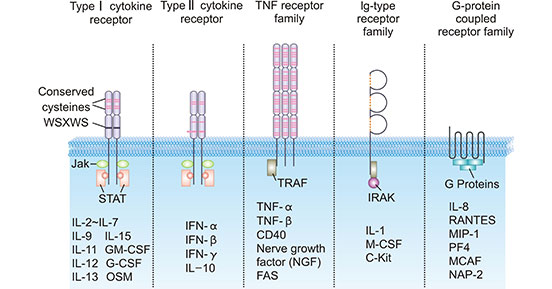
图1. 细胞因子受体的五种典型结构 [1]
*图片来源于Biochim Biophys Acta的出版物
免疫球蛋白(Ig)超家族
免疫球蛋白超家族(IgSF)是指分子结构中具有与免疫球蛋白相似域的分子超家族。IgSF的所有成员都含有1~7个Ig样结构域,每个Ig样结构域含有约70~110个氨基酸残基。它的二级结构是由两条反平行β-折叠状链形成的反平行β-片状平面,每条反平行β-片状链含有3~5个反平行β-折叠。每条反平行β片链由5~10个氨基酸残基组成。β片内侧的疏水氨基酸可稳定Ig的折叠。
大多数Ig域有一个二硫键垂直连接两个β片,构成二硫键的两个半胱氨酸约含55~75个氨基酸。少数Ig域,如CD2的第一域、LFA-3和PDGFR的第四域、CD4的第三域等,均缺乏二硫键。这种多肽链的球形结构的折叠称为免疫球蛋白折叠(Ig fold)。
I型细胞因子受体
I型细胞因子受体又称造血素受体,是表达在细胞表面的跨膜受体,能识别细胞因子并对其作出反应,具有4条α-螺旋链。这些受体具有某些保守的胞外域,缺乏内在的蛋白酪氨酸激酶活性。
保守的胞外域有大约200个氨基酸的长度,其中在氨基末端区域含有四个位置保守的半胱氨酸残基和一个位于跨膜域近端的保守氨基酸基团(WSXWS)。这四个半胱氨酸是维持受体结构和功能完整性的关键。WSXWS共识序列是细胞因子受体功能性蛋白与蛋白相互作用的识别位点。
II型细胞因子受体
II型细胞因子受体又称IFN受体,是表达在某些细胞表面的跨膜蛋白,它与一组选定的细胞因子结合并作出反应。通常Ⅱ型细胞因子受体是具有高亲和力和低亲和力成分的异二聚体或多聚体。这些受体一般由两条肽链组成,胞外区由200个氨基酸残基组成,并含有4个不连续的半胱氨酸。
TNFR超级家族
肿瘤坏死因子受体(TNFR)超家族成员是细胞因子受体的一个蛋白质超家族,共享一个半胱氨酸丰富域(CRD),由三个二硫键围绕CXXCXXC的核心基团形成一个拉长的分子。目前TNFR家族有12个成员,包括55kDa和75kDa的TNFR,低亲和力的NGFR,人B细胞抗原(CD40)和Fas抗原。该家族的共同特点是其胞外区有Cys(4-6)丰富的假重复基团,每个基团含有40个氨基酸残基。细胞内域较短,由44~221个氨基酸残基组成,无同源序列。
趋化因子受体
趋化因子受体是在某些细胞表面发现并与趋化因子相互作用的细胞因子受体。人类已发现20种不同趋化因子受体,为7次跨膜的G蛋白偶联受体,并在细胞内与G蛋白偶联进行信号转导,是G蛋白偶联受体家族成员之一。趋化因子受体与相应的配体结合后,引发细胞内钙(Ca2+)离子通量(钙信号传导)。既而引起细胞反应,包括趋化作用过程开始,将细胞运送到生物体内的理想位置。
参考文献:
[1] Turner MD, Nedjai B, Hurst T, Pennington DJ. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim Biophys Acta. 2014 Nov; 1843 (11):2563-2582. doi: 10.1016/j.bbamcr.2014.05.014. Epub 2014 Jun 2. PMID: 24892271.





