LY6G6D(LY6-G):淋巴抗原6新成员,结直肠癌CRC双抗靶点或免疫老化新指标!
日期:2023-03-17 08:59:56
近期,全球知名生物技术公司基因泰克(Genentech)开发了一种新型LY6G6D/CD3 T细胞依赖性双抗(LY6G6D-TDB),靶向LY6G6D,用于治疗结直肠癌CRC。临床前数据结果表明,LY6G6D-TDB作为单一药物对治疗LY6G6D高表达的结直肠癌CRC具有抗肿瘤功效,当LY6G6D-TDB与PD-1联合用药时可增强疗效。在实体瘤ADC药物治疗中,当正常组织也会表达目标靶标时,正常细胞便会摄入ADC药物,导致“脱靶”毒性效应。因此,LY6G6D在CRC中的特异性高表达是合适临床的理想ADC靶标。
目前广泛报道的结直肠癌靶点包括KRAS、HER2、EGFR、VEGF和GPA33。LY6G6D作为CRC新的治疗靶标,现有相关报道非常少,针对LY6G6D靶点临床药开发,目前公开披露的仅有LY6G6D-TDB。此外,有研究揭示LY6G6D或是免疫衰老功能损害的新标记物。最为重要的是,LY6G6D作为淋巴抗原6超家族(Ly-6SF)新成员,将为CRC双抗治疗注入新的生命力!
1. 什么是Ly6超家族(Ly-6SF)?
Ly6/uPAR/CD59/netuotoxin超家族(Ly-6SF)被称作淋巴抗原6或蛇毒素超家族 [1-2]。1977年,淋巴抗原6(Ly6)首次被报道为一组小鼠淋巴细胞同种抗原。1986年,Ly-6E.1 cDNA首次从小鼠中克隆,并定位在小鼠15号染色体,该位点处还包含其它多基因簇,包括Ly6-A、Ly6-C、Ly-6ThB、Ly6-F、Ly6-G、Ly6-H和Ly-6M,被定义为淋巴抗原6超家族,简称Ly-6SF [1-2]。Ly-6SF大部分为糖基磷脂酰肌醇(GPI)蛋白,通过GPI将蛋白锚定在细胞膜外表面,参与细胞的跨膜信号转导 [1-2]。
目前,至少发现有35种人和61种小鼠Ly6蛋白,如表达在人类肺、肾、心脏、肝脏的CD59;小鼠角质细胞的Lynx1;SLUPR-1则表达在大多数免疫细胞,支气管表皮细胞;CD45,CD43,Sca-1/Ly-6A.2/Ly-6E.1及CD117都表达在小鼠骨髓中。另外,还有银环蛇毒素Cbt、金环蛇毒素aBgt、半环扁尾蛇aEbt等神经毒素。现有研究表明Ly6超家族主要参与免疫调节、信号转导、神经调节和细胞组织发育调节 [3-5]。
2. 什么是LY6G6D?
2.1 LY6G6D的结构
淋巴细胞抗原6复合位点G6D(Lymphocyte antigen 6 complex,locus G6D,LY6G6D,又名LY6-D;LY6-G;G6D;NG25;MEGT1;C6orf23)是淋巴抗原6超家族(Ly-6SF)中的成员之一 [6-7]。LY6G6D基因位于6号染色体上的主要组织相容性复合体(MHC)III类区域,编码133个氨基酸残基的蛋白质,分子量约为13.7 kDa。Ly-6SF蛋白质在结构上有一个显著性的特点,含有一个或多个空间结构保守的Ly6/uPAR(LU)结构域,即以亮氨酸或异亮氨酸为N端开始,包含8-10个半胱氨酸构成的四至五对保守的二硫键,形成三指蛋白结构域(three-fingered protein domain,TFPD),家族成员具有序列保守性低,进化速度快,成员数量庞大并且功能多样化等特点(图1) [7-9]。
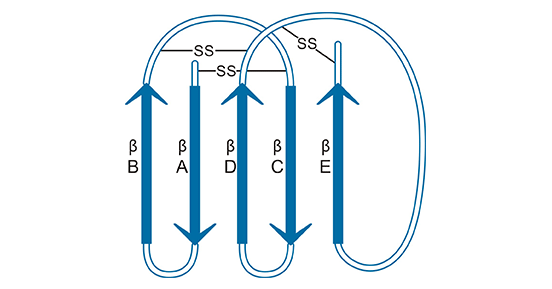
图1. Ly-6SF“三指”蛋白结构示意图 [9]
2.2 LY6G6D的表达
LY6G6D在髓系细胞包括单核细胞、巨噬细胞、粒细胞和中性粒细胞的发育过程中均有不同表达量 [7]。髓系细胞是构成先天免疫反应的主要部分。据报道,CD11b+Ly6G-髓系细胞介导机械性炎症疼痛超敏反应 [10]。LY6G6D通常在单核细胞发育过程中瞬时表达,而在成熟粒细胞和外周血中性粒细胞保留表达,研究表明,LY6G6D在中性粒细胞浸润,招募和迁移中均发挥重要作用 [7]。
3. LY6G6D及家族蛋白相关的调节机制
LY6G6D作为Ly-6超家族的新兴靶点,尽管其机制相关报道十分有限,但最近研究揭示,LY6G6D可能是结直肠癌CRC发生和免疫逃避的关键靶点。LY6G6D高表达增加了治疗后CRC转移性复发的风险。STAT5敲除,显著降低LY6G6D水平,表明STAT5能调控LY6G6D表达。进一步验证发现,JAK/STAT抑制剂可通过靶向STAT5/LY6G6D轴,增加CRC细胞死亡 [11]。
Ly-6家族数量庞大,其结构上较低的保守性也造成了它们功能极具多样性的特点,如C4.4A/LYPD3是在鳞状细胞癌亚型上高表达的一种癌症相关细胞表面蛋白 [12];Lypd6则是存在于Wnt/β-catenin信号通路中的一个正向调节因子,其通过Wnt8调节中胚层和神经外胚层 [13];Lynx1与烟碱型乙酰胆碱受体(nAChR)相互作用,被认为可作为nAChR活性的调节剂发挥作用,以防止过度兴奋,对神经元起到保护作用 [14]。在肺炎感染中,LY6G表达增强,可诱导脾脏自然杀伤细胞中的CD69和KRG1分子被激活 [15, 16];Ly6e被报道参与调节冠状病毒、SARS-CoV、MERS-CoV和SARS-CoV-2的病毒感染 [17];CD59可调控补体激活调节因子MAC,进而参与补体调节的细胞凋亡 [18];CD46能保护宿主不受噬菌体攻击,协同眼睛中的β-1整联蛋白(ITG-Beta1/CD29)相互作用,后者起到视网膜色素细胞粘附作用 [19]。
4. LY6G6D在结直肠癌及免疫老化中的作用
LY6G6D作为Ly-6超家族成员之一,其被鉴定后,国内外相关方面的报道很少,多年来功能不明。然而,最近一些研究开始集中研究LY6G6D和结肠癌发生和发展之间的关系。另有报道还揭示LY6G6D与衰老现象存在密切联系。
4.1 LY6G6D和结直肠癌
结直肠癌(CRC)是胃肠道中常见的恶性肿瘤,然而病因目前尚未明确,在全球范围内结直肠癌已经是最常见的癌症之一。有研究揭示,同正常结直肠组织相比,LY6G6D在原发性和转移性结直肠肿瘤组织中高表达。LY6G6D的高表达与免疫抑制型细胞的高度浸润相关,在CD8+T淋巴细胞减少的结肠癌中,LY6G6D表达和STAT5活化增强。此外,LY6G6D阳性的CRC细胞可通过骨髓源性抑制细胞(MDSC)的扩增,从而抑制T细胞增殖 [20]。
同样在CRC中,有报道显示,p38α MAPK的抑制剂以及DNA甲基转移酶DNMT1敲低,可导致LY6G6D表达降低。在转移性CRC组中,LY6G6D高甲基化,预示着FOLFOX(药物亚叶酸(LV)、氟尿嘧啶(5-FU)和奥沙利铂)一线治疗的耐药性 [21]。因此,在结直肠癌中,LY6G6D差异性高表达与癌症发生密切相关,有望作为结直肠癌潜在预测指标或治疗靶点。
4.2 LY6G6D和免疫老化
免疫系统也会发生衰老,即免疫衰老或免疫老化。衰老是不可避免的生理事件,会影响包括免疫在内的各种生物过程。老化环境中的T细胞容易转变为衰老状态,上调共抑制分子的表达,如PD-1/CD279,TIM-3/CD366,VISTA,其表达变化可导致T细胞功能受损,从而导致整个免疫系统受到干扰。同样地,当中性粒细胞和巨噬细胞等先天免疫细胞的功能发生障碍,可导致病原体入侵,增加慢性炎症和动脉粥样硬化的风险 [22-24]。
一项研究揭示LY6G6D下调是中性粒细胞免疫衰老发生功能损伤的重要指标。实验发现,与年轻老鼠相比,老年小鼠骨髓中性粒细胞中LY6G6D的表达显著下调,并且发现LY6G6D表达水平与中性粒细胞抗细菌免疫中的免疫功能之间存在强相关性,这些免疫能力随着衰老而减弱,如吞噬作用、活性氧(ROS)产生、白细胞介素IL-1β产生、中性粒细胞胞外陷阱(NET)的形成、迁移以及细菌清除 [8]。免疫衰老是导致先天免疫系统严重功能受损的关键事件。然而,目前对骨髓谱系细胞的免疫老化决定开关的机制知之甚少。因而,研究者认为LY6G6D下调可以作为预测中性粒细胞中与年龄相关的功能损害的新标记物。
5. LY6G6D的临床应用前景
多项研究表明,LY-6成员对于调节免疫、神经系统和补体的功能非常重要。特别是在癌症中,众多LY-6成员在许多类型的癌症,例如胃癌、宫颈癌、乳腺癌、卵巢癌、肺癌和膀胱癌中发挥着关键的作用。因此,LY-6基因家族或将在临床实践中发挥重要作用,不仅是疾病预后的一种标志物,而且也是开发新药的重要靶标。深入研究它们的生物学功能,对阐明Ly-6家族功能、分析其结构中蛋白相互作用情况、探索疾病发病机制、发现新的治疗靶点等,将具有重要意义。LY6G6D作为LY-6基因家族新成员,陆续研究报道LY6G6D在结直肠癌等疾病发生过程中扮有重要的角色,如前所述,已有一款基于LY6G6D的双抗药用于结直肠癌治疗,这预示着LY6G6D有望成为基于抗体治疗的一个有潜力的靶点。
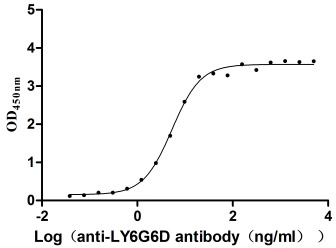
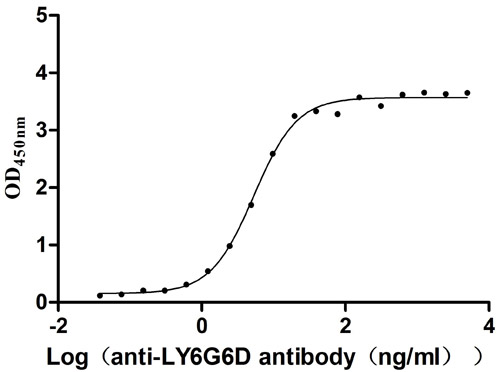
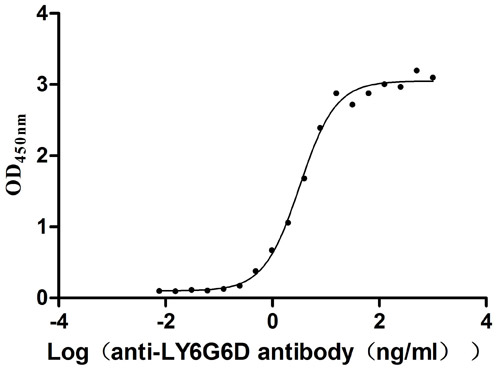
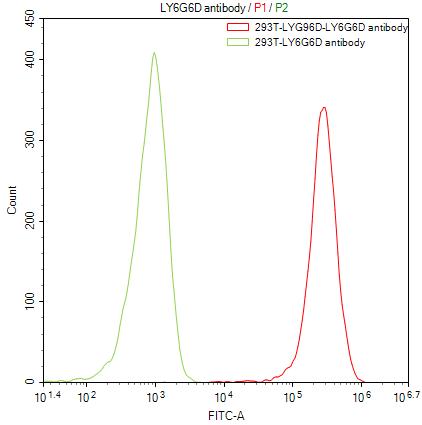
为鼎力协助科研和药企人员针对LY6G6D在机体功能调节方面,以及LY6G6D在结直肠癌和免疫老化相关的临床应用研究,CUSABIO推出LY6G6D活性蛋白(CSB-YP013246HU;CSB-YP4607MOV)产品,助力您在LY6G6D机制方面的研究或其潜在临床价值的探索(点击查看LY6G6D全系列产品:蛋白;抗体)。
LY6G6D Protein
参考文献:
[1] Pflugh, David L., Stephen E. Maher, and Alfred LM Bothwell. "Ly-6I, a new member of the murine Ly-6 superfamily with a distinct pattern of expression. "The Journal of Immunology 165.1 (2000): 313-321.
[2] Guo, Quanyang, et al. "Identification and expression of an uncharacterized Ly-6 gene cluster in zebrafish Danio rerio." Functional & integrative genomics 15 (2015): 577-585.
[3] Rathbun, Luke, Anthony Magliocco, and Anil K. Bamezai. "Upregulated Ly-6 gene expression is associated with poor overall survival in uterine corpus endometrial carcinoma patients." The Journal of Immunology 208.1_Supplement (2022): 179-05.
[4] Swamynathan, Sudha, et al. "Secreted Ly-6/uPAR-related protein-1 (SLURP1) is a pro-differentiation factor that stalls G1-S transition during corneal epithelial cell cycle progression." The Ocular Surface 24 (2022): 1-11.
[5] Bamezai, Anil K., and Julie M. Miwa. "Biology of Ly-6 Supergene Family in Health and Disease. "Frontiers in Cell and Developmental Biology 10 (2022).
[6] Upadhyay, Geeta. "Emerging role of lymphocyte antigen-6 family of genes in cancer and immune cells. "Frontiers in immunology 10 (2019): 819.
[7] Beliakov, I. S., T. A. Karakasheva, and N. N. Mazurenko. "Exon-intron structure of the LY6G6D gene. "Molecular biology 43 (2009): 543-551.
[8] Saito, Suguru, et al. "Lymphocyte antigen 6 complex locus G6D downregulation is a novel parameter for functional impairment of neutrophils in aged mice. " Frontiers in Immunology 13 (2022).
[9] Lee, Pui Y., et al. "Ly6 family proteins in neutrophil biology." journal of leukocyte biology 94.4 (2013): 585-594.
[10] Tan, Peng, et al. "Myeloid loss of Beclin 1 promotes PD-L1 hi precursor B cell lymphoma development." The Journal of clinical investigation 129.12 (2022) : 5261-5277. : 5261-5277.
[11] Caruso, Francesca Pia, et al. "Lymphocyte antigen 6G6D-mediated modulation through p38α MAPK and DNA methylation in colorectal cancer." Cancer Cell International 22.1 (2022): 1-13.
[12] Willuda, Joerg, et al. "Preclinical anti-tumor efficacy of an anti-C4. 4a (LYPD3) antibody drug conjugate for the treatment of lung squamous cell carcinoma." Cancer Research 74.19_Supplement (2014): 5445-5445.
[13] Özhan, Günes, et al. "Lypd6 enhances Wnt/β-catenin signaling by promoting Lrp6 phosphorylation in raft plasma membrane domains." developmental cell 26.4 (2013): 331-345.
[14] Miwa, Julie M. "Lynx1 prototoxins: critical accessory proteins of neuronal nicotinic acetylcholine receptors." current opinion in pharmacology 56 ( 2021): 46-51.
[15] Broquet, Alexis, et al. "Depletion of natural killer cells increases mice susceptibility in a Pseudomonas aeruginosa pneumonia model." critical care medicine 42.6 (2014): e441-e450.
[16] Wang, Wenxiu, et al. "Exogenous interleukin-33 promotes hepatocellular carcinoma growth by remodelling the tumour microenvironment." journal of translational medicine 18 (2020): 1-15.
[17] Pfaender, Stephanie, et al. "LY6E impairs coronavirus fusion and confers immune control of viral disease." Nature microbiology 5.11 (2020): 1330-1339.
[18] Farkas, Imre, et al. "CD59 blocks not only the insertion of C9 into MAC but inhibits ion channel formation by homologous C5b-8 as well as C5b -9." The Journal of physiology 539.2 (2002): 537-545.
[19] McLaughlin, Barbara J., et al. "Novel role for a complement regulatory protein (CD46) in retinal pigment epithelial adhesion. "Investigative ophthalmology & visual science 44.8 (2003): 3669-3674.
[20] Giordano, Guido, et al. "JAK/Stat5-mediated subtype-specific lymphocyte antigen 6 complex, locus G6D (LY6G6D) expression drives mismatch repair proficient colorectal cancer." Journal of Experimental & Clinical Cancer Research 38.1 (2019): 1-11.
[21] Caruso, Francesca Pia, et al. "LY6G6D is Epigenetically Activated in Classical Colorectal Adenocarcinoma and Hyper-Methylated in Mucinous Subtypes Determining Resistance to FOLFOX Therapeutic Regimens."(2021).
[22] Batista-Duharte, Alexander, et al. "Immune Checkpoint Inhibitors for Vaccine Improvements: Current Status and New Approaches." Pharmaceutics 14.8 ( 2022): 1721.
[23] Zych, Michał, et al. "Differences in Immune Checkpoints Expression (TIM-3 and PD-1) on T Cells in Women with Recurrent Miscarriages- Preliminary Studies." Journal of Clinical Medicine 10.18 (2021): 4182.
[24] Liu, Jun, et al. "Immune-checkpoint proteins VISTA and PD-1 nonredundantly regulate murine T-cell responses. "Proceedings of the National Academy of Sciences 112.21 (2015): 6682-6687.










