趋化因子受体
趋化因子(chemokine)是一种分子量为8~10 kDa的多肽,是最大的细胞因子家族, 主要功能是招募血液中的单核细胞、中性粒细胞、淋巴细胞等进入特定的淋巴器官和组织以及感染发生的部位。趋化因子受体是一类介导趋化因子行使功能的七次跨膜G蛋白偶联受体(GPCR),通常表达于免疫细胞、中性粒细胞、内皮细胞等细胞膜上。分子由约330个氨基酸组成。趋化因子及趋化因子受体在介导细胞迁移、增殖和抵御病原体入侵过程中发挥重要作用,并且与免疫环境中炎症和癌症的发生发展密切相关。最近,趋化因子受体比趋化因子本身受到更多关注,一方面是它们具有明显的结构特征,另一方面是趋化因子受体的缺乏现在与一些免疫缺陷状态直接相关。
1、趋化因子结构与功能
趋化因子受体是一种7次跨膜的G蛋白偶联受体。GPCR的结构生物学研究一直充满挑战,它们在异源系统中表达产量很低,也很难从中提取纯化目的蛋白并保持其稳定性。此外,由于涉及多次跨膜,如何表达出有活性且构象完整的全长多次跨膜蛋白一直是困扰GPCR结构和功能研究的一大难题。针对多次跨膜蛋白,j9九游会登录入口首页生物搭建了病毒样颗粒 (VLPs)、去垢剂和Nanodisc三大技术平台,极大程度上解决了全长跨膜蛋白制备难的问题(了解更多>>)。
趋化因子受体的7个跨膜区将分子分成细胞外自由N-端、3个细胞外环、3个细胞内环和C-端几个部分(如图1所示)。胞内第二环是与异三聚体G-蛋白偶联的部位,有特征的天门冬氨酸-精氨酸-酪氨酸盒(DRYbox)氨基酸序列。与趋化因子受体偶联的异三聚体G-蛋白的α亚基为Gi/o,对百日咳毒素敏感。
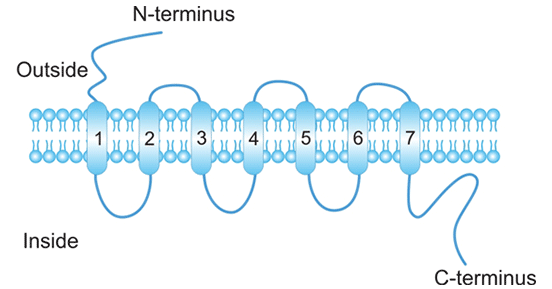
图1. 趋化因子受体结构示意图
趋化因子受体被趋化因子等配体激活后,主要通过结合胞内的G蛋白或阻遏蛋白(β-arrestin)等信号分子来进行下游信号转导。趋化因子及其受体在病原体的清除、炎症反应、病原体感染、细胞及器官的发育、创伤的修复、肿瘤的形成及其转移等方面都起着重要的作用。
2、趋化因子受体类型
趋化因子受体通常分为两大类:典型趋化因子受体(cCKR)和非典型趋化因子受体(ACKR)。
目前有18个cCKR,根据它们结合的主要趋化因子类型(即CC、CXC、CX3C或XC)命名,后跟字母 R(表示“受体”),阿拉伯数字反映的是它们被发现的顺序。非典型趋化因子受体(有四种)在结构上与 cCKR 相关,但不与cCKR 激活的信号转导通路偶联。这些成员如下:
● 典型趋化因子受体 (cCKRs)
● 非典型趋化因子受体(ACKRs)
3、趋化因子受体信号
趋化因子受体的细胞内信号传导依赖于以 (αβγ) 异源三聚体形式存在的G蛋白。它们由三个不同的亚基组成。Gα亚基直接与趋化因子受体细胞内环和Gβ亚基相互作用,后者又与Gγ亚基形成紧密复合物。Gα亚基包含一个参与GTP结合和水解的GTPase 结构域。当分子GDP 与G蛋白亚基结合时,G蛋白处于非活性状态。当趋化因子配体与趋化因子受体结合时,它会诱导趋化因子受体形成激活细胞内异源三聚体G蛋白的构象,从而导致GDP与GTP的分子交换。然后Gα-GTP从受体和Gβγ异二聚体解离,Gα亚基激活与细胞膜相关的磷脂酶C (PLC)。PLC裂解磷脂酰肌醇(4,5)-二磷酸(PIP2)形成两个称为三磷酸肌醇(IP3)和二酰基甘油(DAG)的第二信使分子;DAG 激活另一种称为蛋白激酶 C (PKC) 的酶,IP3触发细胞内储存的钙释放。这些事件促进了许多信号级联反应,从而影响细胞反应。您可以单击此处查看信号通路。
4、趋化因子受体与疾病
在过去的几十年里,趋化因子及趋化因子受体因在人体细胞迁移和抵御病原体入侵中起到的重要作用而引起了人们的广泛关注。除了它们在HIV病理机制方面的作用外,人们也更多地认识到它们在自身免疫及炎症、癌症方面的重要影响。尤其是癌症方面,趋化因子及其受体的表达在致瘤性中起双重作用。一方面,由癌症起始细胞或它们周围正常细胞分泌的趋化因子可以通过增加白细胞向该部位的迁移来限制肿瘤的发展,并诱导长期的抗肿瘤免疫。另一方面,它们可能促进肿瘤细胞的存活、增殖和转移潜能。恶性细胞中最常过度表达的趋化因子受体是 CXCR4。它存在于超过23种不同类型的人类癌症中,例如肺癌、脑癌、前列腺癌、乳腺癌、胰腺癌、卵巢癌、结直肠癌、白血病和黑色素瘤。恶性细胞上的 CXCR4 表达与细胞存活、肿瘤生长、血管生成、更高的转移潜能和对治疗剂的抗性相关。它的配体CXCL12由骨髓、淋巴结、肝和肺细胞大量分泌。随着单克隆技术的发展,针对CXCR4、CCR2、CCR8和CCR4的单克隆抗体已经进入癌症治疗的临床试验。





